اصلاح سنتی در مقابل اصلاح مدرن:
سه دوره اصلی اصلاح نباتات را می توان شناسایی کرد: (1) اصلاح نباتات بر اساس انتخاب گونه های مشاهده شده، بدون توجه به منشاء آنها. (ب) تولید و انتخاب تنوع گسترش یافته توسط تلاقی کنترل شده. و (iii) نظارت بر وراثت تنوع درون ژنوم و انتخاب نوترکیبهای خاص. چهارمین مرحله اصلاح نباتات را می توان ایجاد و معرفی تنوع جدید به ژنوم ها از طریق مهندسی ژنتیک دانست.
- الف) اصلاح نباتات بر پایه ی تنوع مشاهده شده
1- اهلی کردن گیاهان (منشاء گیاهان)
2- انتخاب شهودی توسط کشاورزان (منشاء نژادهای محلی : Landrace)
3- انتخاب لاین های خالص و انتخاب توده ای ( منشاء کولتیوارها)
- ب) اصلاح نباتات بر پایه ی تلاقی های کنترل شده
1- اصلاح شجره ای ( بازی با والدین)
2- اصلاح آیدوتایپ (بازی با صفات)
3- اصلاح جمعیت (بازی با تنوع ژنتیکی)
4- اصلاح هیبرید (بازی با هتروزسیس)
- ج) اصلاح نباتات بر پایه ی مونیتورینگ نوترکیبی
1- نقشه یابی ژنها مورد توجه ( یافتن سوزن در انبار کاه)
2- انتخاب بر پایه ی نشانگرها (MAS): ساخت ژنوتیپهای متناسب
3- انتخاب ژنوم: سرعت بخشیدن به پیشرفت ژنتیکی
الف) اصلاح نباتات بر پایه ی تنوع مشاهده شده
ابتدایی ترین شکل اصلاح نباتات انتخاب گونه های طبیعی در طبیعت و بعدها در مزارع کشت بود. تنوع ژنتیکی به طور مداوم تحت فشار انتخاب چرخه جمع آوری غذا یا کاشت - برداشت قرار می گیرد. در برخی موارد، این فرآیند منجر به تغییرات عمیق در فنوتیپهای گیاهی میشود که نمونه آن استخراج ذرت از تئوسینت است. این مرحله اولیه اصلاح نباتات از زمان پیدایش کشاورزی تا اولین آزمایش های هیبریداسیون که توسط کلروتر در دهه 1760 انجام شد را در بر می گیرد. با کشف قوانین وراثت، در نوبت از قرن 19 به قرن 20، اهمیت هیبریداسیون در اصلاح گیاهان به طور گسترده ای شناخته شد. امروزه تقریباً تمام برنامههای اصلاح نباتات شامل استفاده از هیبریداسیون است.
1- اهلی کردن گیاهان (منشاء گیاهان)
برای یک ژن معین، جهشها رویدادهای نادری هستند، اما با توجه به تعداد زیاد گیاهان در یک مزرعه و ژنهای موجود در یک گیاه، جهشها رویدادهای بسیار مکرر در یک جمعیت هستند. بیشتر جهشها برای بقا در طبیعت نامطلوب هستند و در چند نسل در نتیجه انتخاب طبیعی از جمعیت حذف میشوند. با این حال، برخی از این جهش ها ممکن است منجر به فنوتیپ های مطلوب تری از نظر کشت یا از نظر کیفیت غذا شود. برخی از آن جهشیافتهها توسط کشاورزان باستانی نجات یافتند، که از آنها در برابر رقابت محافظت کردند و با گیاهانی که در غیر این صورت ناتوان بودند رابطه همزیستی برقرار کردند. برخلاف زیستگاههای وحشی، مزارع کشتشده محیطهایی بودند که در آن جهشها مزیت انتخابی ایجاد میکردند، بنابراین از طریق انتخاب انسان به نوع غالب تبدیل شدند. تجمع این نوع جهش عامل اصلی سندرم اهلیسازی است، مجموعهای از ویژگیها که بسیاری از گونههای کشتشده را برای بقای خود به طور غیرقابل برگشتی به انسان وابسته میکند.
تنوع مولکولی در گیاهان اهلی کمتر از گونههای وحشی مرتبط است، به عنوان یک نتیجه از اثر بنیانگذار در طول اهلیسازی. با انتخاب شدید گیاهان جهش یافته نادر سازگار با کشت، کشاورزان اولیه بیشتر تنوع موجود در جمعیتهای وحشی را که اشکال کشت شده از آنها به وجود آمدند، حذف کردند. اکنون مشخص شده است که بسیاری از ژنهای ارزشمند، بهویژه آنهایی که مربوط به مقاومت در برابر آفات هستند، از مخزن ژنی کشتشده کنار گذاشته شدهاند. ترکیب آن ژنها در ارقام مدرن، بدون از دست دادن ز
مین از نظر عملکرد و کیفیت محصول، یکی از چالش های اصلاح نباتات مدرن و یکی از مرتبط ترین کاربردهای ابزار مولکولی در برنامه های اصلاحی است.
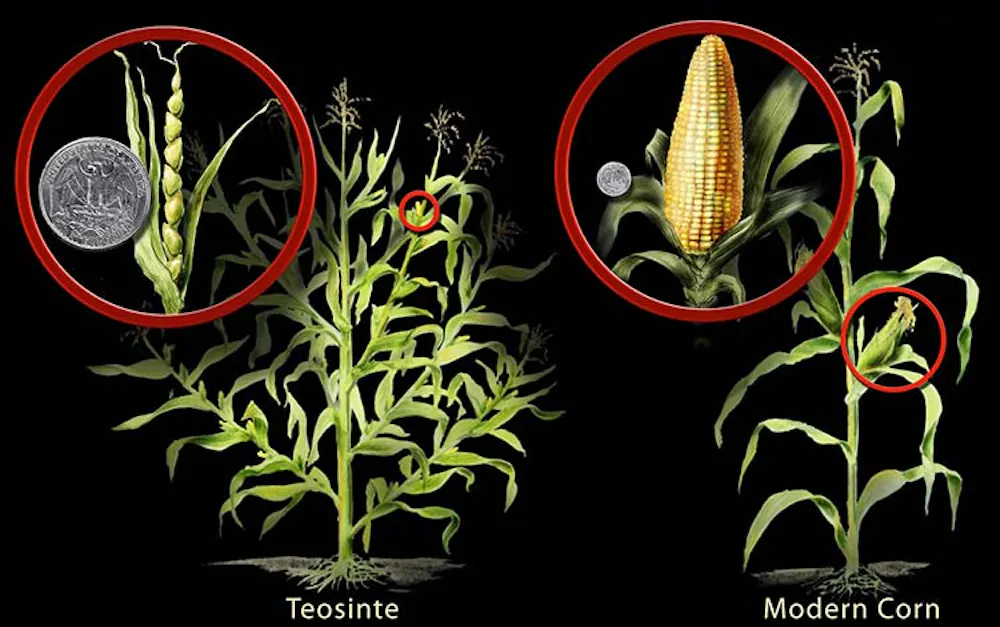
2- انتخاب شهودی توسط کشاورزان (منشاء نژادهای بومی: Landrace)
نژادهای بومی جمعیتهایی از گیاهان هستند که برای نسلهای زیادی در یک منطقه خاص کشت شدهاند و تحت تأثیر تنشهای زیستی و غیرزیستی، مدیریت محصول، نگهداری بذر و ترجیحات غذایی شکل میگیرند. آنها موجودات ژنتیکی پویا هستند: به طور مداوم در نتیجه انتخاب عمدی و غیرعمدی، مخلوط بذر و تبادل گرده در حال تغییر هستند. نژادهای بومی با تعادل بین انتخاب تثبیت کننده، که هویت نژاد بومی را در یک منطقه معین حفظ می کند، و انتخاب جهت ملایم شکل می گیرند که منجر به تعدیل آهسته تغییرات محیطی می شود. در برخی موارد، تغییرات سریع ممکن است رخ دهد، به خصوص زمانی که نژاد بومی به منطقه دیگری منتقل می شود یا زمانی که مواد جدید در نزدیکی بومی اصلی کشت می شوند. اگر تولید بذر تایید شده متوقف شود و بذرهای ذخیره شده توسط کشاورز به طور مکرر، بدون مراقبت برای جداسازی در برابر آلودگی بذر یا گرده، کاشته شوند، امروزه هنوز هم می توانند از ارقام مدرن مشتق شوند.
ویژگیهای اصلی نژادهای بومی عبارتند از (1) سطوح بالای تنوع ژنتیکی در جمعیتها، که با طیف محدودی از تنوع بین افراد مشخص میشود، با ویژگیهای متمایزی که نژاد بومی را قابل شناسایی میکند. (2) سازگاری با شرایط خاک و آب و هوای معمول منطقه، همراه با مقاومت در برابر آفات رایج؛ (3) بخشهای خوراکی که توسط مردم محلی ارزشمند هستند و معمولاً توسط غذاهای محلی شکل میگیرند. و (4) عملکرد متوسط اما پایدار، امنیت غذایی به جامعه محلی تحت تغییرات طبیعی محیطی اعطا می کند.
انتخاب شهودی کشاورز این مزیت را دارد که تنوع را برای محیط واقعی و خاص مورد استفاده و ترجیحات غذایی محلی شکل میدهد، که به خوبی در مورد کشاورزی معیشتی (Subsistence agriculture)، جایی که بیشتر تولید به صورت محلی مصرف میشود، خدمت میکند. با این حال، هنگامی که کشاورزان برای یک صفت انتخاب می کنند، همبستگی ژنتیکی ممکن است منجر به تغییرات نامطلوب در سایر صفات شود. به عنوان مثال، تودههای غلات معمولاً گیاهان بلندی هستند، مستعد ورس (Lodging) و نشان دادن شاخص برداشت پایین، که دلیل احتمالی آن نتیجه ی انتخاب انسان برای قسمتهای خوراکی بزرگ ( خوشه، بلال، خوشه) است.
با این وجود، نژادهای بومی به دلیل تنوع ژنتیکی و سازگاری با محیطهای مختلف، ارزشمندترین منابع ژنتیکی برای برنامههای بلندمدت اصلاح نباتات و همچنین اهداف اصلی برای تهیه ی مجموعههای ژرم پلاسم هستند. بانک های بذر در سراسر جهان هزاران نمونه از نژادهای بومی را تحت حفاظت "ex situ" نگهداری می کنند. در برخی کشورها، تلاشهایی برای ابداع مکانیسمهای نظارتی و مشوقهای مالی برای جوامع سنتی وجود دارد تا به رشد انواع میراث خود ادامه دهند، و هدف آن حفاظت «in situ» است. سیستمهای جدیدی برای حفاظت از ژرم پلاسم در شبکههای اجتماعی ساخته شدهاند که افراد علاقهمند به این موضوع را به عنوان یک سرگرمی به یکدیگر متصل میکنند (مانند seedsavers.org). این شبکه ها از ابزارهای ارتباطی مدرن بهره می برند تا آنچه را که قبلاً از طریق تماس شخصی در جوامع سنتی اتفاق می افتاد در مقیاس جهانی تکرار کنند.
با وجود این، بخش بزرگی از تنوعی که زمانی در مزارع کشت گیاهان یکساله وجود داشت، ممکن است در طی معرفی ارقام مدرن و پرمحصول به طور غیر قابل برگشتی از بین رفته باشد. از این نظر، همان ارقام مدرنی که میلیونها نفر را از گرسنگی نجات دادند، ممکن است گونههایی را که حاصل قرنها انتخاب شهودی محلی توسط کشاورزان و منبع ارزشمندی برای بهبود ژنتیکی آینده بودند، از بین ببرند.
3- انتخاب لاین های خالص و انتخاب توده ای ( منشاء کولتیوارها)
اولین روش اصلاح نباتات مبتنی بر دانش ابتدایی قوانین وراثت، انتخاب گیاهان در داخل نژادهای بومی بوده است، بر اساس این فرض که انتظار می رود نتاج بهترین افراد نسبت به نسلی که از آن نمونه تصادفی انتخاب شده، برتری داشته باشند. جمعیت این روش به طور رسمی توسط لوئیس دو ویلمورین در سال 1856 پیشنهاد شد، اگرچه اشاره هایی به استفاده از اصول آن توسط برخی کشاورزان در اوایل قرن 19 وجود دارد. این درک را می توان منشأ الگوی همگنی دانست که تا به امروز بر اصلاح نژاد و کشاورزی در کل حاکم است. از این نقطه به بعد، ناهمگونی درون مزرعه ای نامطلوب در نظر گرفته شد و هم اصلاح نباتات و هم زراعت روش هایی را برای دستیابی به حداکثر همگنی فضایی (به عنوان مثال، "کشاورزی دقیق"؛ Precision agriculture) توسعه دادند.
در گونه های خود گرده افشان مانند برنج و گندم، نژادهای بومی را می توان مخلوطی از لاینهای خالص در نظر گرفت، از جمله برخی افراد هتروزیگوت که از فرکانس پایین دگر گرده افشانی به دست می آیند. در این نوع جمعیت، انتخاب گیاهان منفرد و مشتق کردن نتاج اینبرد همواره منجر به ایجاد برخی از لاینهایی میشود که در شرایط رشد معین از نژاد اصلی بهتر عمل میکنند. با این حال، این برتری هزینه دارد، زیرا خطوط خالص معمولاً نسبت به جمعیتهای متنوع در مواجهه با استرسها، بهویژه بیماریها، پایداری کمتری دارند و ظرفیت سازگاری طولانیمدت را ندارند، زیرا برای اکثر ژنها یک شکل هستند.
در مورد گونههای با گردهافشانی باز، مانند ذرت، نژادهای بومی جمعیتهایی از افراد حاصل از جفتگیری تصادفی هستند که تقریباً تعادل هاردی-واینبرگ (HWE) را با برخی انحرافات به دلیل انتخاب خفیف تشکیل میدهند. انتخاب انبوه و بازترکیب بخشی از جمعیت با عملکرد برتر منجر به افزایش تدریجی فراوانی آللهای مطلوب میشود. نسلهای متوالی انتخاب در نژادهای بومی ذرت منجر به بهبود واریتههای گردهافشانی شده شد که تا ظهور ذرت هیبریدی اساس تولید ذرت بود.
ب) اصلاح نباتات بر پایه ی تلاقی های کنترل شده
علیرغم تنوع خود به خودی بزرگی که می توان در نژادهای بومی یافت، به کارگیری ساده انتخاب بر روی تنوع موجود، یک فرآیند فرسایشی است که در نهایت به محدودیت می رسد. قدرت خلاق واقعی اصلاح نباتات در ترویج نوترکیبی برای به هم زدن آلل های مطلوب است. ترکیب آلل های مختلف در بسیاری از جایگاه ها منجر به تعداد بی نهایت ژنوتیپ می شود. می توان یک برنامه اصلاح نژاد تجاری را از ده ها والدین بنیانگذار و سازگار با تمرکز واضح بر یک محیط هدف خاص و ارزیابی نتاج حجیم در حال تفرق آغاز کرد. تزریق تنوع جدید ممکن است در مورد تغییر قابل توجه محیط هدف، مانند ظهور آفات جدید که مواد پایهدهنده مقاومتی برای آنها نداشتند، ضروری باشد.
با توجه به تعداد بیشماری از ژنوتیپهای احتمالی ناشی از تلاقی والدین متنوع، محدودیت برای دستاوردهای ژنتیکی به ظرفیت برنامه اصلاحی برای ارزیابی تعداد زیادی از گیاهان، حاصل از تعداد زیادی تلاقی تبدیل میشود. به همین دلیل، اصلاح نباتات اغلب بازی اعداد نامیده میشود و برنامههای رقابتی بزرگ در کالاها سرمایهگذاری زیادی بر روی روشهای بازده بالا برای جابجایی، کاشت، ارزیابی و برداشت بذر میکنند. با انباشته شدن دستاوردهای ژنتیکی، این نوار به تدریج بالا می رود و سرمایه گذاری های فزاینده ای بیشتری برای حفظ نرخ ثابت پیشرفت ژنتیکی مورد نیاز است. حد این تشدید، قابلیت مالی بازده در بازار بذر و تجارت مرتبط است. روشهای اصلی توسعه یافته برای استفاده کارآمد از منابع در برنامه های اصلاحی در ادامه مورد بحث قرار می گیرد.
1- اصلاح شجره ای ( بازی با والدین)
اکثریت قریب به اتفاق ارقام آزاد سازی شده از گونه های خود گرده افشان از طریق روش شجره ای توسعه یافته اند. اصلاح شجره شامل تلاقی والدین و تولید جمعیت هایتفرق یافته است که از طریق نسل های خود گرده افشانی و انتخاب انجام می شود تا زمانی که مجموعه ای از لاین مشتق شده که ویژگی های خوب هر دو والدین را ترکیب می کند به دست آید. از آنجایی که این روش مبتنی بر تکمیل صفات است، این روش برای اصلاح صفات کیفی مانند مقاومت به بیماری یا صفات به راحتی قابل طبقه بندی مانند معماری گیاه یا رنگ یا شکل اجزای گیاه کارآمد است. روش شجره نامه برایاصلاح کنندگان جذاب است زیرا امکان ساخت واریته های بهتر را با کنار هم قرار دادن صفات خوب موجود در مواد مختلف در یک گیاه فراهم می کند. از آنجایی که تمام تلاقی ها کنترل شده است، می توان از شجره نامه هر رقم مطلع شد.
ضعف اصلی روش شجره نامه در این واقعیت است که عملکرد تنها در پایان فرآیند، در لاین های اینبرد، زمانی که بذر برای آزمایش های تکراری در دسترس است، به طور موثر ارزیابی می شود. اگرچه در این مرحله، مگر اینکه تعداد زیادی از لاینها پیشرفت کرده باشند، فضای کمی برای بهبود پتانسیل بازده وجود دارد. در نتیجه، نرخ پیشرفت عملکرد حاصل از روش شجره نامه معمولاً متوسط است و به ندرت از 1٪ در سال تجاوز می کند.

2- اصلاح آیدوتایپ (بازی با صفات)
رویکرد اصلاح ایدئوتیپ (Ideotype breeding) را می توان به عنوان یک استراتژی برای بهبود ظرفیت روش شجره نامه در جهت ارتقاء سود برای صفات کمی، به ویژه عملکرد در نظر گرفت. بر اساس این فرضیه است که می توان صفات پیچیده را با تغییر صفات ساده تر که همبستگی مثبت با آنها دارند، بهبود بخشید. مزیت این روش این است که در صورت صحت فرضیه زیربنایی، می توان سود قابل توجهی را برای عملکرد، حتی با یک برنامه اصلاحی کوچک، با اتخاذ رویکرد "هوشمندانه"، بر خلاف رویکرد "بازی اعداد" افزایش داد. برنامه های اصلاحی در مقیاس بزرگ علاوه بر این، از نظر علمی برای اصلاح کنندگان جذاب است، زیرا آنها شانس تغییر الگوها (paradigms) در محصول مورد علاقه خود را دارند. با این حال، مهم است که به خاطر داشته باشید که همبستگیهای ژنتیکی نامطلوب میتواند مزیتهای حاصل از صفات تشکیلدهنده ایدئوتیپ را جبران کند.
تغییر ایدئوتیپ یک محصول اغلب مستلزم جستجوی تغییرات فراتر از مرزهای ژرم پلاسم الیت (elit) است که معمولاً از نوع گیاخان فعلی است. با این حال، استفاده از نژادهای بومی به عنوان والدین در برنامه های اصلاحی معمولاً منجر به کاهش قابل توجه عملکرد می شود. به همین دلیل، بک کراس به سمت مواد الیت برای بازیابی نتاج رقابتی ضروری است. اگر دهنده صفت جدید یک خویشاوند وحشی باشد، مشکلات سازگاری جنسی، عقیمی گل، کیفیت دانه یا پراکندگی دانه (یعنی خرد شدن دانه) ممکن است برای چندین نسل باقی بماند.
برای جلوگیری از اثرات نامطلوب ورود مستقیم مواد عجیب و غریب به جمعیتهای اصلاحکننده الیت، این مواد معمولاً ابتدا در مرحله پیشزادآوری (prebreeding) استفاده میشوند، زمانی که اصلاحکنندگان سعی میکنند ارتباط بین صفات مفید و نامطلوب را از بین ببرند. هنگامی که صفت جدید مورد نیاز برای جمع آوری ایدئوتیپ در یک پس زمینه الیت درج شد، آن لاینهای انتقالی را می توان به برنامه پرورش الیت منتقل کرد.

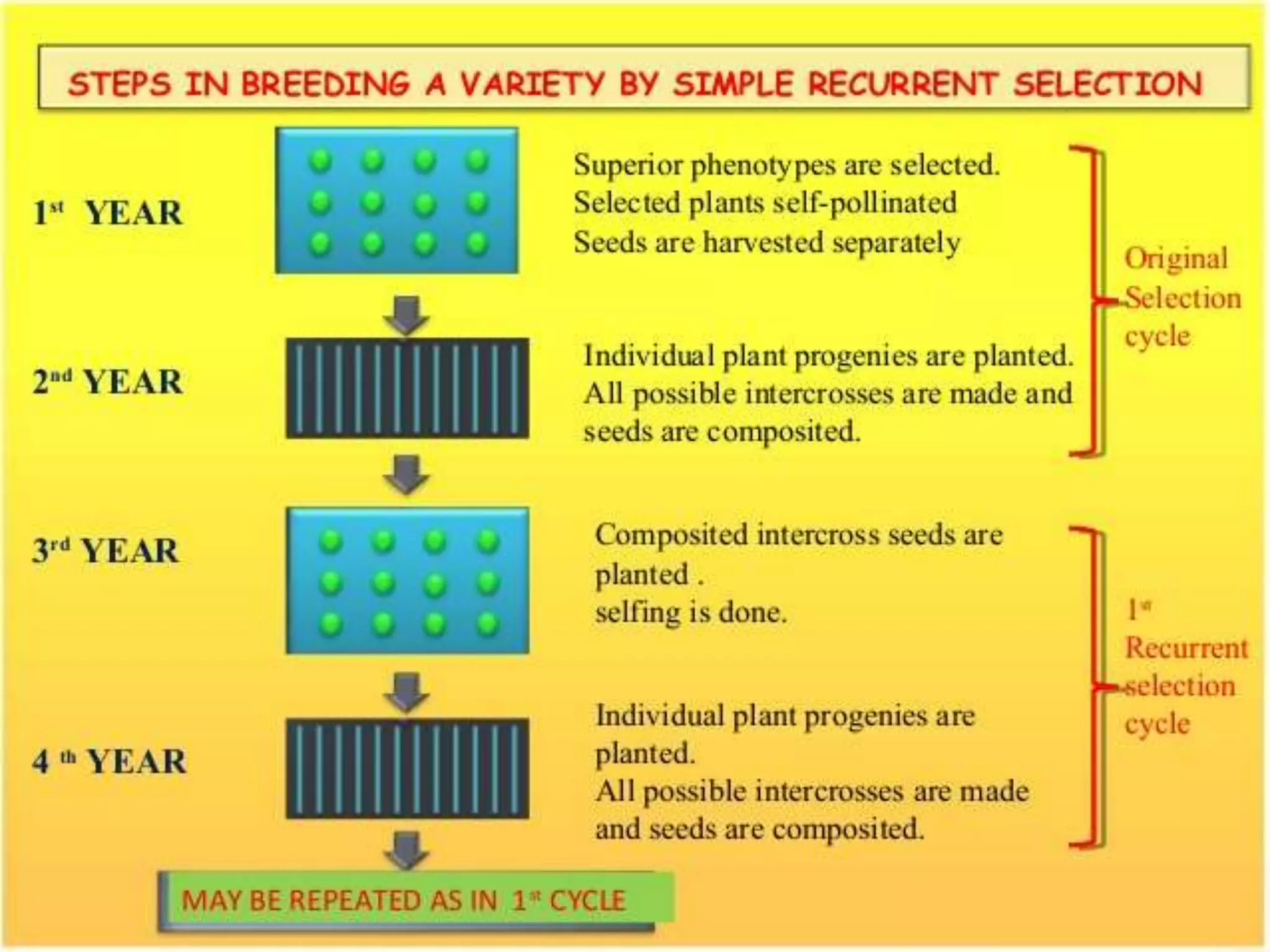
3- اصلاح جمعیت (بازی با تنوع ژنتیکی)
اگرچه همه روشهای اصلاحی بر اعمال فشار انتخاب بر روی یک جمعیت متغیر دلالت میکنند، اما اصطلاح «اصلاح جمعیت» (population breeding) روشی را نشان میدهد که برای بهبود عملکرد فنوتیپی یک جمعیت در هم آمیخته با افزایش فراوانی آللهای مطلوب کنترل کننده ی صفات مورد توجه طراحی شده است. ساده ترین نسخه اصلاح جمعیت، روش انتخابتوده ای است که برای گونه های دگر گرده افشان اعمال می شود، که در آن جمعیت بهبود یافته به طور مستقیم به عنوان یک رقم استفاده می شود. بعدها، طرح های پیچیده تری برای اصلاح جمعیت طراحی شده است که چارچوبی را برای توسعه نظریه ژنتیک کمی فراهم می کند. در اصلاح جمعیت مدرن، هدف افزایش ارزش جمعیت به عنوان منبعی از لاین های الیت است. بهبود میانگین کیفیت جمعیت، در عین حفظ تنوع در آن، منجر به ایجاد افراد برتر می شود که از لاین قبلی بهتر عمل می کنند. این لاین ها را می توان به عنوان ارقام در مورد گونه های خودگرده افشان یا به عنوان والدین هیبریدها در مورد گونه های دگرگرده افشان استفاده کرد.
اصلاح جمعیت یک طرح باز از دورهای متوالی انتخاب و نوترکیبی است، بنابراین به عنوان اصلاح انتخاب دوره ای (recurrent selection breeding) نیز شناخته می شود. انتخاب دوره ای نیاز به یک سیستم تلاقی کارآمد دارد که می تواند محدودیتی برای گونه های خود گرده افشان باشد. این طرح، هنگامی که به طور مکرر بر روی جمعیت های کوچک اعمال می شود، منجر به کاهش تنوع ژنتیکی و کاهش دستاوردهای ژنتیکی می شود. به همین دلیل، یک جمعیت موثر بزرگ باید در سراسر جهان حفظ شود، در غیر این صورت رانش ژنتیکی بر سایر نیروهایی که بر جمعیت اثر میگذارند، غلبه میکند و مانع پیشرفت ژنتیکی میشود.
با این حال، حتی اندازه متوسط جمعیت (به عنوان مثال، 50 گیاه) برای جلوگیری از کاهش سریع تنوع ژنتیکی کافی است. این احتمالاً به دلیل این واقعیت است که در حالی که انتخاب منجر به از بین رفتن تنوع در برخی از ژن ها می شود، نوترکیبی بلوک های لینکاژی را به قطعات کوچکتر می شکند و امکان ترکیب ژن های بسیار بیشتری را نسبت به قطعات بزرگ DNA فراهم می کند. این پدیده تغییرات پنهان را در نسلهای بعدی در فرآیند انتخاب مکرر آزاد میکند و از دست دادن تنوع ناشی از انتخاب را جبران میکند.
اصلاحگران با تلاقی چندین لاین که به عنوان منابع آلل های مطلوب برای یک یا چند صفت انتخاب می شوند، جمعیت های جدیدی ایجاد می کنند. این جمعیتهای مصنوعی تحت انتخاب مکرر، رویدادهای ژنتیکی را تقلید میکنند که قبلاً در نژادهای بومی در دست کشاورزان سنتی رخ میداد، با این تفاوت که کل فرآیند نظارت و کنترل میشود و فشار انتخاب برای دستیابی سریعتر تشدید میشود. نرخ بهره ژنتیکی در واحد زمان را می توان با سرعت بخشیدن به چرخه های انتخاب-نوترکیب، با تشدید فشار انتخاب، با بهبود دقت ارزیابی (در نتیجه افزایش وراثت پذیری)، یا با هر ترکیبی از آنها افزایش داد.
طرح کلی اصلاح جمعیت بسیار منعطف است و امکان سفارشی سازی برای نیازها و اهداف خاص گونه های مختلف و برنامه های اصلاحی را فراهم می کند. این سیستم را میتوان برای نتایج سریع و کوتاهمدت، معمولاً با اعمال فشار انتخاب قوی بر روی جمعیتهای مبتنی بر پایه ی ژنتیکی محدود، یا برای نتایج پایدار و طولانیمدت، با اعمال فشار انتخاب متوسط بر روی جمعیتهای مبتنی بر پایه ی ژنتیکی گسترده، قالببندی کرد. اصلاح جمعیت همچنین می تواند به عنوان یک طرح پیش از اصلاح (Prebreeding) مورد استفاده قرار گیرد، زیرا رویدادهای متقابل دوره ای آن باعث ایجاد نوترکیبی بین ژنوم های بیگانه (exotic) و الیت می شود و ژن های بیگانه نامطلوب را از جمعیت پاک می کند.
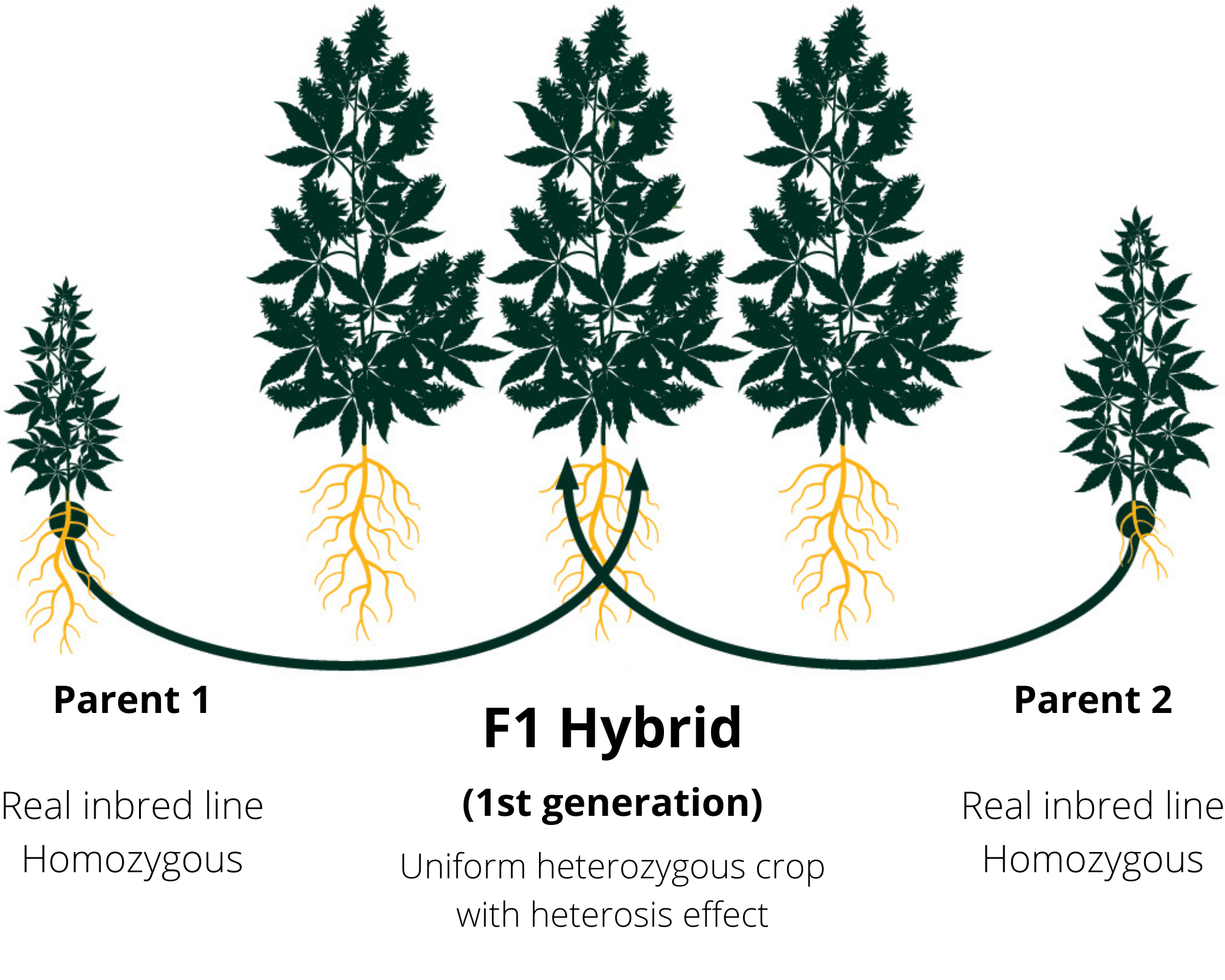
4- اصلاح هیبرید (بازی با هتروزسیس)
هتروزیس برتری افراد هیبرید نسبت به افراد اینبرد است. در حدود معینی، هر چه والدین واگراتر باشند، هتروزیس در فرزندان آنها بیشتر است. بنیه هیبرید به سرعت در طول نسل های اینبرد کاهش می یابد، که نشان می دهد، مکانیسم اصلی هتروزیس هر چه باشد، به دلیل وجود جایگاه های هتروزیگوت است. به همین دلیل، امروزه برنامه های پرورش ذرت بر روی توسعه هیبریدهای رقابتی F1 متمرکز شده است که در آنها هتروزیگوز در حداکثر مقدار خود است.
دو چالش در برنامه های اصلاح نژاد هیبرید وجود دارد: (1) نیاز به بهبود حداقل دو جمعیت به سمت سازگاری زراعی، در حالی که فاصله ژنتیکی آنها به اندازه کافی برای بیان هتروزیس قوی است، و (2) توسعه تولید کارآمد بذر هیبریدهای منتخب، به طوری که هزینه تولید بذر ارزش عملکرد اضافی ناشی از هتروزیس را جبران نمی کند. در ذرت، اولین مشکل به مفهوم گروههای هتروتیک منجر شد، که مخزن ژن الیت را به زیرمجموعههایی تقسیم کرد، که در آن اصلاح جمعیت اعمال میشود. مشکل دوم با بهبود لاینهای والدینی که منجر به عملکرد بیشتر در مزارع تولید بذر شد و با استفاده از ژنهای نر عقیمی و اخته کردن مکانیکی ("detasselling") برای جلوگیری از خود گرده افشان حل شد.
از سوی دیگر، هیبریدها از منظر تجاری مزایای زیادی دارند. بذرهای تولید شده توسط گیاهان هیبرید به دلیل تفکیک هزاران ژن از نظر ژنتیکی ناهمگن هستند. در صورت کاشت، محصول حاصل تنوع زیادی در صفات زراعی، معماری گیاه و طول چرخه ایجاد می کند و در نتیجه عملکرد و کیفیت دانه کاهش می یابد. از این رو، کشاورزان باید هر سال بذرهای جدید بخرند و در نتیجه تقاضای دائمی برای بذر هیبریدی وجود دارد. علاوه بر این، با ظهور تراریخته ها، دانه های هیبریدی به حامل های ترجیحی آن صفات اختصاصی ارزشمند تبدیل شدند، زیرا امکان کنترل بهتر رویداد توسط مالک را فراهم می کنند.
ج) اصلاح نباتات بر پایه ی مونیتورینگ نوترکیبی
روش های پرورش سنتی بر اساس مکمل بودن بین ویژگی های والدین بود. با این حال، اطلاعات کمی در مورد اینکه کدام بخش از ژنوم از هر یک از والدین گرفته شده است، وجود نداشت. این وضعیت با ظهور و انتشار فناوریهای نشانگر مولکولی تغییر کرد که امکان نظارت بر انتقال بخشهای کروموزوم در نتاج را فراهم کرد.
تقریباً از هر گونه تغییر در توالی بین افراد می توان برای طراحی نشانگری استفاده کرد که امکان شناسایی والدینی را که در بخش خاصی از کروموزوم در یک لاین نوترکیب نقش داشته است، می دهد. تا همین اواخر، محبوب ترین نشانگرها تکرارهای توالی ساده (SSR) بودند که به عنوان ریزماهواره ها نیز شناخته می شدند. این نشانگرها توسط پلیمورفیسمهای تک نوکلئوتیدی (SNPs) جایگزین شدند که در ژنوم فراوانتر هستند و بیشتر در معرض ژنوتیپسازی با توان عملیاتی بالا هستند.
نشانگرهای مولکولی ابزارهای ضروری برای مطالعه کنترل ژنتیکی هر صفت مورد علاقه هستند که در نهایت منجر به شناسایی ژنهای زیربنایی این صفت و زنجیرههای متابولیکی درگیر میشوند. این مکان را می توان به طور گسترده به عنوان زیست شناسی مولکولی تعریف کرد که در دهه های گذشته بر حوزه علوم زیستی تسلط داشت.
1- نقشه یابی ژنها مورد توجه ( یافتن سوزن در انبار کاه)
هنگامی که بسیاری از نشانگرهای مولکولی در مجموعه ای از گیاهان به دست آمده از یک تلاقی منفرد ژنوتایپینگ می شوند، می توان از فراوانی نوترکیبی بین آنها برای استنباط ترتیب و فاصله نسبی آنها در کروموزوم ها استفاده کرد و در نتیجه یک نقشه ژنتیکی به دست آمد. اگر آن گیاهان یا نتاج آنها برای یک صفت کمی ارزیابی شوند، می توان یک مدل آماری ایجاد کرد که در آن بخشی از واریانس فنوتیپی را می توان با برخی از نشانگرها توضیح داد، که به این معنی است که آن نشانگرها باید به ژن های زیربنایی مرتبط باشند. صفت این رویکرد منجر به نقشههای مکان صفت کمی (QTL) میشود که معمولاً اولین گام به سمت درک کنترل ژنتیکی یک صفت کمی است. از نقشه QTL، یک اصلاحگران گیاه می تواند بینشی در مورد تعداد جایگاه های کنترل کننده یک صفت، اهمیت نسبی آنها و موقعیت تقریبی آنها در ژنوم بدست آورد. در مورد QTLهای با جلوه بزرگ، ممکن است با استفاده از نزدیکترین نشانگرهای موجود در نقشه، مستقیماً به انتخاب به کمک نشانگر بروید. با این حال، در بیشتر موارد، یک مرحله از نقشه برداری دقیق، شامل جمعیت بزرگتر و فاصله نشانگر متراکم تر در ناحیه ژنومی هدف، برای توسعه ابزارهای انتخاب مفید ضروری است. نتیجه نهایی این رویکرد، شناسایی ژن و پلی مورفیسم در توالی نوکلئوتیدی آن است که مسئول تفاوتهای فنوتیپی مشاهدهشده است.
رویکرد نقشه برداری QTL برای توضیح تضاد بین دو والدین موثر است، اما برای بررسی تنوع ژنتیکی گسترده تر برای یک صفت در ژرم پلاسم ناکارآمد است. رویکرد نقشه برداری ارتباطی (association mapping) میانبری در این مسیر ارائه می دهد و به همین دلیل جذابیت زیادی برای اصلاحگران دارد. در این رویکرد، پانلی از لاینهای ژنتیکی متنوع به طور متراکم ژنوتایپینگ شده و به دقت برای یک ویژگی فنوتیپی ارزیابی می شود، با هدف شناسایی ارتباط بین آلل های نشانگر و صفات. از آنجایی که فاصله شجره نامه ای بین مواد در پانل زیاد است، این ارتباط تنها در صورتی قابل توجه خواهد بود که نشانگر به طور محکم با ژن عامل پیوسته باشد یا اگر عوامل مرتبط با ژنتیک جمعیت (ساختار جمعیت) بین جایگاه های غیر مرتبط ارتباط ایجاد کند. در مورد دوم، برآورد ساختار جمعیت و در نظر گرفتن صریح آن در مدل آماری میتواند از تشخیص تداعیهای نادرست جلوگیری کند. هنگامی که به درستی اعمال شود، تجزیه و تحلیل ارتباط می تواند منجر به تشخیص نواحی ژنومی مربوط به صفت مورد نظر و به طور همزمان، در شناسایی لاینهای اهداکننده آلل های مطلوب از یک ژرم پلاسم گسترده تر شود.
بانک های ژن هزاران توده را در خود جای داده اند که به طور بالقوه برای اصلاح نباتات مفید هستند. این الحاق ها شامل اقوام محصولات وحشی و توده های منسوخ شده (obsolete landraces) است. اگرچه ارزش زراعی ضعیفی در مقایسه با ارقام مدرن دارند، اما اعتقاد بر این است که این مواد دارای ژنهای مفیدی هستند که از گونههای وحشی در فرآیند اهلیسازی یا از نژادهای بومی در مراحل اولیه اصلاح علمی گرفته نشدهاند. نجات آن ژنهای مفیدی که «رها شدهاند» (left behind) کار دشواری است، زیرا ژنهای مورد نظر ممکن است بهطور محکم با ژنهای نامطلوب پیوسته باشند، که به جمعیت در حال اصلاح کشیده میشوند. درک تنوع ژنتیکی در مجموعه های ژرم پلاسم اولین گام به سمت استفاده بهتر از یک مخزن ژنی گسترده تر در برنامه های اصلاحی است. تلاقی برگشتی به کمک نشانگر امکان شناسایی نوترکیبهای نادر در مجاورت ژن معرفیشده و ویرایش آن بخش از کروموزوم را تا حد ممکن نزدیک به ژن هدف را فراهم میکند. علاوه بر این، با شناسایی افراد دارای تمیزترین ژنوم (با کمترین باقیمانده از والد اهداکننده)، می توان تعداد دفعات تلاقی برگشتی مورد نیاز برای بازیابی کامل فنوتیپ الیت را کاهش داد.
مجموعهای از لاینهای با قطعات کوروموزمی جایگزین شده (کتابخانههای CSSL: chromosome segment substitution lines ) مجموعهای از لاینهای مشتقشده از یک تنوع الیت هستند که در آن هر لاین دارای یک بخش از کروموزوم است که با بخش مربوطه در گونههای وحشی مورد علاقه جایگزین میشود. کتابخانه CSSL خرد شده ی کل ژنوم خویشاوند وحشی را در انبارها حمل می کند. اثر فنوتیپی مجموعهای از ژنها در هر انبار را میتوان در برابر لاین الیت اصلی که به عنوان یک بررسی استفاده میشود، ارزیابی کرد. این ذخایر ژنتیکی میتوانند شناسایی و ورود ژنها از خویشاوندان وحشی به ژرم پلاسم الیت را تسهیل کنند. CSSL های برتر را می توان برای نقشه برداری دقیق و توسعه همزمان یک نسخه بهبود یافته از انواع الیتها استفاده کرد.
2- انتخاب بر پایه ی نشانگرها (MAS): ساخت ژنوتیپهای متناسب
در حالی که در مورد ورود ژن به کمک نشانگر،اصلاحگران به طور معمول تنها یک ژن را برچسب گذاری می کند، یک طرح انتخابی با کمک نشانگر با ویژگی های کامل، چندین ژن را به طور همزمان بررسی می کند. آگاهی از آلل حامل هر گیاه در هر مکان اصلی مورد علاقه، ابزاری را برای ساخت ترکیبات آللی خاص ایجاد می کند که ارزش زراعی لاین را به حداکثر می رساند. با این حال، هنگامی که با چندین ژن سروکار داریم، مهم است که در نظر داشته باشیم که تعامل بین ژن ها (اثر اپیستاتیک) و نه تنها اثرات افزایشی ژن ها، بیان یک صفت را مشخص می کند.
یک کاربرد معمولی از انتخاب به کمک نشانگر، هرم سازی ژن مقاومت (resistance gene pyramiding) است. هنگامی که فردی با بیماریهای ناشی از پاتوژنهایی سروکار دارد که تنوع بالایی دارند، یک ژن مقاومت ممکن است برای چند نسل گیاهی مقاومت کامل ایجاد کند تا زمانی که توسط سویههای پاتوژن بر آن غلبه شود. تجمع چندین ژن مقاومت در انواع مختلف می تواند مقاومت بادوام (durable resistance) ایجاد کند، زیرا پاتوژن باید همه ژن ها را به طور همزمان شکست دهد، که به شدت احتمال شکست مقاومت را کاهش می دهد. هرم کردن ژن مقاومت با روشهای اصلاحی کلاسیک تقریباً غیرممکن است، زیرا تأثیر یک ژن تنها در غیاب سایر ژنهای مقاومت قابل مشاهده است. ژن های مقاومت با برچسب نشانگر را می توان به راحتی بدون نیاز به غربالگری فنوتیپی ترکیب کرد. یک استراتژی مشابه را می توان برای رشد پیشگیرانه مقاومت به پاتوژن هایی که در حال حاضر در یک منطقه وجود ندارند استفاده کرد، زیرا در غیاب پاتوژن ها هیچ غربالگری مقاومت فنوتیپی امکان پذیر نخواهد بود.
نشانگرهای مولکولی مورد علاقه برای پرورش دهندگان یک گونه معین را می توان در مجموعه های بهینه برای ژنوتیپ همزمان ("مولتی پلکس": multiplexed) ترکیب کرد، که امکان استفاده با توان بالا را در برنامه های اصلاحی کاربردی، به ویژه آنهایی که با یک پایه ژنتیکی گسترده تر سروکار دارند، را می دهد. اجرای چنین مجموعه نشانگری به حجم وسیعی از تحقیقات قبلی بستگی دارد. برای هر یک از صفات مورد علاقه، چندین مرحله تحقیق مورد نیاز است، که معمولاً با نقشه یابی مکان های صفت کمی (QTL) شروع می شود، سپس نقشه برداری دقیق و در نهایت نقشه برداری موقعیتی (fine mapping) انجام می شود. هنگامی که ژن عامل بر اساس یک جفت آلل متضاد شناسایی شد، کاوش آللهای جدید در ژرم پلاسمهای متنوع ممکن است دامنه تنوع صفت یا جعبه ابزار را برای مقابله با تنشهای خاص گسترش دهد.
3- انتخاب ژنوم: سرعت بخشیدن به پیشرفت ژنتیکی
پیشرفتهای اخیر در فناوری ژنوتیپسازی به شدت هزینه ژنوتیپسازی را کاهش داد و امکان امتیاز دهی هزاران نشانگر در جمعیتهای گیاهان تحت انتخاب را ایجاد کرد. چندین مرحله از تجزیه و تحلیل ژنتیکی برای شناسایی رابطه هر یک از آن نشانگرها با فنوتیپ نمی تواند با آن همگام باشد مورد نیاز است. در این سناریو، اصلاحکننده میتواند مقدار زیادی اطلاعات ژنوتیپی با رابطه نامعلوم با صفات انتخابی داشته باشد. رویکرد انتخاب ژنومی پیشنهاد میکند که آگاهی از رابطه بین نشانگرهای خاص و ژنهای خاص در زمینه اصلاحی ضروری نیست. در عوض، اصلاحکننده میتواند از اطلاعات موجود برای همه نشانگرهای یک گیاه برای پیشبینی ارزش اصلاحی آن استفاده کند، بدون اینکه فنوتیپ آن را ارزیابی کند، بر اساس مدلهای آماری قبلی ساخته شده برای آن نشانگرها با استفاده از «جمعیت آموزشی» (training population) که همه ژنوتیپها و فنوتیپها برای آن تعیین شدهاند. گل زد. نشانگرها به عنوان عوامل تصادفی در چارچوب تحلیل مدل مختلط در نظر گرفته میشوند، زیرا تعداد نشانگرهای مورد استفاده در انتخاب ژنومی معمولاً بر تعداد افراد در جامعه آموزشی برتری دارد، به طوری که تخمین اثر هر نشانگر ممکن نخواهد بود. به فقدان درجات آزادی مطالعات شبیه سازی نشان داد که انتخاب ژنومی می تواند پیشرفت ژنتیکی صفات کمی در اصلاح جمعیت را تسریع کند.
انتخاب ژنومی می تواند به عنوان روشی برای به حداکثر رساندن بازده مرحله ارزیابی فنوتیپی در چرخه اصلاحی دیده شود. اطلاعات به دست آمده از یک دور ارزیابی فنوتیپی مناسب می تواند به نسل های بعدی انتشار یابد. هنگامی که مدلها برای محیطهای مختلف ساخته میشوند، پروفایل منحصربهفرد نشانگر برای یک فرد معین میتواند برای پیشبینی عملکرد آن در هر یک از این محیطها مورد استفاده قرار گیرد، که استتنتاج انجامشده روی عملکرد هر فرد را به طور قابلتوجهی بهبود میبخشد و هزینههای ارزیابی را تا حد زیادی کاهش میدهد. استفاده بالقوه از این رویکرد در انتخاب والدین جمعیتهای اصلاحی جدید نیز امیدوارکننده است. با این حال، پیشبینیپذیری فنوتیپ از پروفایل نشانگر در طول نسلها محو میشود و دورهای جدیدی از دادههای فنوتیپی باید برای بهروزرسانی مدل آماری وارد شوند. این رویکرد به ویژه در مواردی مفید خواهد بود که ارزیابی فنوتیپی دشوار، پرهزینه، یا مشروط به رویدادهای نامشخص، مانند وقوع یک آفت یا شرایط آب و هوایی خاص باشد.
انتخاب ژنومی می تواند برای تسریع پرورش برای پتانسیل عملکرد مورد استفاده قرار گیرد، که اغلب به عنوان سخت ترین صفت برای انتخاب به کمک نشانگر در نظر گرفته می شود. در صنایع بذر بسیار رقابتی، مانند موارد ذرت و سویا، هر گونه افزایش در نرخ بهره ژنتیکی می تواند تفاوت قابل توجهی در سهم بازار ایجاد کند. به همین دلیل، شرکتهای بزرگ بذر سرمایهگذاری زیادی در آنالیز DNA با توان عملیاتی بالا و بیوانفورماتیک برای ادغام یکپارچه انتخاب ژنومی در برنامههای اصلاحی انجام میدهند. دانه ها را می توان قبل از کاشت ژنوتایپینگ کرد، به طوری که فقط افراد از پیش انتخاب شده در کرتهای گران قیمت در مزرعه استفاده می کنند. با توجه به میزان سرمایه گذاری مورد نیاز برای تنظیم و به روز نگه داشتن لاین کشی ژنوتیپ، در مقایسه با شرکت های بذر کوچک، موسسات دولتی و دانشگاه ها، کاربرد کامل انتخاب ژنومی باید در شرکت های خصوصی بزرگ آسان تر اتفاق بیفتد، چرخش زمانی برای تجزیه و تحلیل DNA، پردازش و انتخاب داده ها و تیم آموزش دیده مورد نیاز است.
Author
Alexandre Siqueira Guedes Coelho - Universidade Federal de Goiás, Escola de Agronomia e Engenharia de Alimentos, Campus Samambaia, Goiânia, GO, Brazil 74690-900
اصلاح نباتات تکاملی (Evolutionary Plant Breeding)
انتخاب طبیعی و سایر فرآیندهای تکاملی در اصلاح نباتات تکاملی به منظور ایجاد گونههای گیاهی جدید استفاده میشوند. هدف این استراتژی تکرار فرآیندهای طبیعی به منظور ایجاد گیاهانی با ویژگیهای مطلوب، مانند مقاومت در برابر بیماری یا عملکرد بالا است. کار با گونههای موروثی یا گیاهان وحشی، که تنوع ژنتیکی بیشتری نسبت به بسیاری از محصولات کشاورزی تجاری دارند، یک روش معمول در اصلاح نباتات تکاملی است.
پرورش دهندگان می توانند با استفاده از تنوع طبیعی در این جمعیت های گیاهی، گونه های جدیدی ایجاد کنند که به خوبی با شرایط رشد منطقه ای سازگار بوده و دارای کیفیت های ارزشمند متنوعی هستند.
اصلاح گیاهان زینتی (Ornamental Plant Breeding )
هدف از پرورش گیاهان زینتی ایجاد گیاهانی با ویژگی های جذاب یا تزئینی، مانند گل های پر جنب و جوش، برگ های جذاب، یا اشکال متمایز است. در محوطه سازی، باغبانی و سایر اهداف تزئینی، این گیاهان اغلب برای جذابیت زیبایی شناختی خود پرورش داده می شوند.
پیش اصلاحی (Prebreeding):
Pre-breeding refers to all activities designed to iden- tify desirable characteristics and/or genes from unadapted materials that cannot be used directly in breeding populations and to transfer these traits to an intermediate set of materials that breeders can use further in producing new varieties for farmers
قبل از اصلاح به تمام فعالیتهایی اطلاق میشود که برای شناسایی ویژگیها و/یا ژنهای مطلوب از مواد ناسازگاری که نمیتوانند مستقیماً در جمعیتهای اصلاحی استفاده شوند و برای انتقال این صفات به مجموعهای میانی از مواد طراحی شدهاند که اصلاحگران میتوانند بیشتر در تولید انواع جدید واریته های زراعی
1- انتخاب جهت دار به نفع افراد برجسته در جمعیت؛ این نوع انتخاب به نفع افراد با ارزش های صفت در یکی از افراطی های توزیع صفت عمل می کند
2- نسل اول گیاهان هیبرید F1 که از تلاقی بین دو لاین هم نژاد مشتق شده است
جمعیتهای اثر بنیانگذار که از تعداد کمی از افراد نشات میگیرند، به دلیل خطای نمونهگیری از آللها، به طور قابلتوجهی با جمعیتی که آن افراد از آن سرچشمه گرفتهاند، متفاوت هستند.
رانش ژنتیکی تغییرات ناخواسته در فراوانی های آللی به دلیل واریانس نمونه برداری ذاتی نمونه های کوچک
شرایط تئوریک تعادل هاردی-واینبرگ (HWE) توسط جمعیت های پانمیتی نامتناهی که تحت جهش، مهاجرت یا انتخاب نیستند به دست آمده است. تحت HWE، فرکانس های هموزیگوت ها و هتروزیگوت ها مواردی هستند که به طور تصادفی انتظار می رود، که صرفاً تابعی از فرکانس های آللی هستند.
نسبت شاخص برداشت بین توده دانه و زیست توده کل، به صورت تک بوته یا تاج پوشش
گروه هتروتیک از ژنوتیپ ها که توانایی ترکیبی و پاسخ هتروتیک مشابهی را هنگام تلاقی با ژنوتیپ های سایر گروه های ژرم پلاسم نشان می دهند.
پدیده اثر پلیوتروپیک نسبت به یک ژن که بیش از یک صفت را تحت تأثیر قرار می دهد
تفاوتهای ساختار ژنتیکی جمعیت در ساختار ژنتیکی زیرجمعیتهایی که بخشی از جمعیت مرجع بزرگتر هستند
جایگاه جایگاه صفت کمی در ژنوم که به تعیین یک صفت فنوتیپی با توزیع پیوسته کمک می کند.
صفات کمی که دارای یک تغییر مداوم هستند که معمولاً از تأثیر همزمان بسیاری از ژن ها و محیط ناشی می شود.
تثبیت انتخاب انتخاب به نفع افراد معمولی در جمعیت. این نوع انتخاب علیه افرادی با ارزش های صفت شدید عمل می کند
